Streptococci
definition
The term streptococci is a type of bacteria that share certain common morphological and functional properties.
Most streptococci are harmless and part of the normal human flora. Few of them can cause infections.

What groups of streptococci are there?
Streptococci are divided into a total of three groups. First, a distinction is made between the so-called alpha-hemolytic streptococci and the beta-hemolytic streptococci. The distinction is made based on how they break down the red blood pigment hemoglobin, i.e. how they hemolyze.
The alpha-hemolytic streptococci include the pneumococci and the Viridans pneumoniae, which also belong to the streptococci.
The beta-hemolytic streptococci are further subdivided into the A, B and D streptococci. This further subdivision takes place on the basis of different sugar chains which are embedded in the bacterial wall.
If you're more interested in this topic, then check out our next article: Bacteria
A streptococci
The A-streptococci belong to the beta-hemolytic streptococci. The best-known pathogen of the A streptococci is the Streptococcus pyogenes.
This pathogen causes infections in the area of the nasopharynx in particular. Examples of this are acute tonsillitis (Tonsillar angina), i.e. inflammation of the tonsils, scarlet fever, otitis media or various infections of the soft tissues.
B streptococci
The most prominent pathogen from the group of B streptococci is the Streptococcus agalactiae. It is a typical trigger of meningitis in newborns (Neonatal meningitis) or neonatal sepsis, colloquially known as blood poisoning.
In adults, streptococcus agalactiae can cause wound and bone infections and inflammation of the urethra (Urethritis).
Streptococcus mutans
The Streptococcus mutans belongs to the group of alpha-hemolytic streptococci. It is one of the most important causes of tooth decay.
This pathogen has some special properties which help it to form bacterial plaques on the surface of the tooth. For example, the bacterium can convert carbohydrates into lactic acid. This makes the environment around the tooth more acidic, which is harmful to the tooth substance.
Furthermore, Streptococcus mutans can produce certain proteins that reduce the local immune defense in the mouth so that the body cannot fight the bacterium effectively.
These streptococci are dangerous to humans
Few streptococci can cause infections in humans. How dangerous these are then depends, among other things, on the stage of life and the strength of the immune system.
The human pathogenic streptococci, i.e. those that can trigger a disease in humans, include the pneumococci, the viridans streptococci, Streptococcus pyogenes, Streptococcus agalactiae and the enterococci, which strictly speaking also belong to the streptococci.
These strep infections do exist
The different streptococci can cause very different ranges of infections. Therefore, the most important bacteria and their typical clinical pictures are discussed.
In the group of alpha-hemolytic streptococci are the pneumococci (Streptococcus pneumoniae) the most important representatives. As their name suggests, they like to solve pneumonia (pneumonia) out.
In the event of an infection, however, otitis media, sinusitis or even meningitis can develop.
The second, important representative from this group are the Viridans streptococci. They occur in the oral cavity, intestines and vagina and are often involved in mixed infections such as tooth decay or appendicitis.
Streptococcus A infections typically lead to acute tonsillitis or scarlet fever. In rare cases, soft tissue or bone infections are also possible.
In the case of B streptococci, infection with the Streptococcus agalactiae most significant. For example, the infection can lead to meningitis or blood poisoning in newborns, or inflammation of the urethra. Infections in the middle ear or the salivary gland are rare but possible.
Do you have any further interest in this topic? Then read our next topic below: Strep infection
These symptoms are how I recognize a strep infection
The symptoms of an infection with streptococci depend very much on which organ is infected with the bacteria. Therefore a general statement is hardly possible.
Pneumococcal pneumonia manifests itself as a high fever and productive cough with yellowish-greenish mucus. In addition, the breathing rate may be increased and a subjective feeling of shortness of breath may be present.
Do you now suspect pneumonia? You can find out more at: Signs of pneumonia
An acute inflammation of the tonsils by A streptococci causes rather unspecific symptoms such as fever, headache, cough or sore throat. A lumpy language, as well as pain when swallowing and the resulting difficulty swallowing, are also typical.
If, after the symptoms listed above, you suspect you may have tonsillitis, read our next article below: Signs of tonsillitis
Scarlet fever is also associated with a fever and difficulty swallowing. In addition, an enlargement of the cervical lymph nodes and the typical strawberry or raspberry tongue are observed in scarlet fever.
Read more about signs of scarlet fever below: The symptoms of scarlet fever
Middle ear infections can be triggered by various streptococci. The signs of an otitis media are particularly noticeable through sudden, severe and one-sided ear pain. In addition, fever, hearing impairment or dizziness can occur.
It is important to note that the symptoms can be atypical in very young children as well as in old people, so that one cannot immediately infer the underlying clinical picture.
Streptococci are so contagious
There is no precise measure of the "infectious power" of bacteria. However, streptococci can spread in a variety of ways, which favors the infection.
How long are streptococci contagious?
If streptococci are treated with antibiotics, they are no longer contagious after about 24 hours. If antibiotic therapy is stopped prematurely or without antibiotics, streptococci can be contagious for up to several weeks.
This is the typical way of infection
The typical route of infection is direct or indirect contact between two people. Therefore, in the event of an infection, a good level of hygiene should be observed and public facilities should only be visited after the infection has subsided.
In some cases, strep can be transmitted to a second person through the air. This is called droplet infection and is, for example, the typical way of infection for streptococcal angina.
This is how streptococci are transmitted
- Through direct contact
Streptococci are typically transmitted through direct contact. Streptococci often get on the hands first and are then passed on to the mucous membranes of the nasopharynx when the face is touched.
But they can also survive on surfaces and thus be indirectly transmitted to a second person.
- By the droplet infection
Another possible transmission path is via the so-called droplet infection. When coughing or sneezing, tiny droplets are released into the air, which can then be inhaled by a second person.
Since the droplets can also contain pathogens such as streptococci, they are carried to the nasal and oral mucosa and can cause an infection there.
The Streptococcus agalactiae is also often transmitted through sexual intercourse in infections of the urethra. It is therefore important to treat both partners with such an infection.
You can read more about this topic under: Droplet infection
We have these streptococci on our skin
A large number of different streptococci are typical pathogens of the skin flora. So they appear on the skin without causing an infection. A distinction must be made between those streptococci, which cause a disease in humans.
The beta-hemolytic streptococci like the Streptococcus pyogenes or agalactiae can cause soft tissue infections on the skin. This happens especially when the skin is very dry and cracked or has already been damaged by athlete's foot, for example.
Read more about this topic under: Strep Rash - How Can I Relieve Symptoms?
Strep in the nose
There is a large number of streptococci that do not cause disease in humans, which means that they are not pathogenic to humans. Many of these harmless streptococci occur naturally on the mucous membrane in the nasopharynx.
Each person harbors a different composition of bacteria on the nasal mucous membrane, which, among other things, can also play important roles. If the nose, especially in the winter months, is infected by human pathogenic streptococci, such as the Streptococcus pneumoniae infected, it comes to the clinical picture of sinusitis.
This is then expressed by a purulent secretion from the nose and a feeling of pressure in the area of the affected sinus. In addition, you may experience a fever, a blocked nose and a general feeling of illness.
The infection usually goes away on its own, so that testing for streptococci often does not have to be carried out and no antibiotic is required at all.
However, if there are risk factors such as immunosuppression or high fever, antibiotic therapy can be considered.
Streptococci in the urine
Ideally, the urine should be sterile. This means that no bacteria should be present in the urine. However, a number of bacteria colonize the genital area, including the area around the exit of the urethra, so that contamination can often occur.
Both the Streptococcus agalactiae as well as the enterococci can cause infections of the urinary bladder or urethra, whereby streptococci can then be found in the urine.
The Streptococcus agalactiae causes inflammation of the urethra (Urethritis). In men, urethritis manifests itself with a burning sensation in the urethra and so-called Bonjour drops. The Bonjour drop is a small drop of pus that can be seen in the morning before urinating for the first time.
In women, a discharge from the vagina and pelvic pain are typical with urethritis. The antibiotic doxycycline is used for treatment, and the partner should also be treated so that there can be no re-countertransference during sexual intercourse.
Enterococci in the urine suggest a bladder infection. Women in particular often suffer from bladder infections due to their short urethra. Typical for the clinical picture are frequent, painful urination and a burning sensation when urinating.
The uncomplicated cystitis can be treated with a single dose of fosfomycin, for example.
You can read more about this topic under: Bacteria in the urine - how dangerous is it?
Streptococci in the vagina
In the vagina, lactic acid bacteria protect against infections with other bacteria, such as streptococci. The lactic acid bacteria, as their name suggests, produce lactic acid and keep the area around the vagina as acidic as possible.
Frequent sexual intercourse with changing partners, regular vaginal rinsing, antibiotic therapies or stress can lead to an imbalance in the mucous membrane flora of the vagina.
Pathogens such as streptococci, but above all the Gardnerella vaginalis can colonize the mucous membrane and thereby trigger the clinical picture of bacterial vaginosis.
Clinically, there is a discharge from the vagina, which is thin to foam and can take on colors such as white, gray or yellow. A fishy odor, which is caused by the breakdown of proteins, is also typical.
In addition, the pH value is also increased, which is due to the low production of lactic acid. Some women also report burning pain during sexual intercourse, itching in the genital area and painful urination.
Any bacterial vaginosis should be given antibiotic therapy to prevent the infection from rising to the uterus.
Read about this topic under: Bacterial Vaginosis - What To Do?
Which antibiotics are best for treating strep throat?
Pneumococci are the most common cause of pneumonia, especially in young adults. Typical symptoms are high fever, purulent sputum and an increased respiratory rate with shortness of breath.
The antibiotic of choice for pneumococcal pneumonia are aminopenicillins, such as amoxicillin.
In addition to pneumococci, pneumonia in newborns can also be caused by streptococcus agalactiae.
For more information on the antibiotic described above, see: Amoxicillin - field of application and effect
These antibiotics help against strep throat
In Germany, most streptococcal strains are still sensitive to penicillins such as penicillin G, ampicillin or amoxicillin. It is also possible to give macrolide antibiotics such as erythromycin or cephalosporins (ceftriaxone or cefuroxime).
The exact dosage and the form of administration depend on the respective clinical picture. Depending on which bacterium caused the infection, there is a different antibiotic of choice. For example, penicillin G is the antibiotic of choice for pneumococcal infection.
This means that if possible, penicillin G should also be given in the event of such an infection and not another broader antibiotic, so that no resistance is cultivated.
In the group of Viridans streptococci, the antibiotic of choice is the cephalosporin ceftriaxone. In the case of enterococci, ampicillin or linezolid is preferred if resistance is suspected.
Read more about this topic under: Treatment with antibiotics
Streptococci in pregnancy - you need to pay attention to this
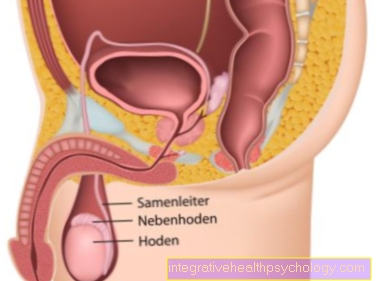
Streptococci that colonize the exit of the vagina and rectum are particularly relevant during pregnancy. These are B streptococci.
If these streptococci are present, they can be passed on to the newborn at birth. There is then a risk that the pathogens can cause meningitis or pneumonia or blood poisoning in the newborn.
To avoid this risk, a swab of the vagina and rectum can be taken between the 35th and 37th week of pregnancy and examined for B-streptococci. If there is actually a colonization, the woman is recommended to take antibiotic prophylaxis.
The smear is a voluntary examination that has to be paid for by yourself. In addition, it is not entirely undisputed and the examination is only being considered for risk groups, for example in women with premature rupture of the bladder or if an earlier child is infected.
During pregnancy it is of course also possible to contract streptococci and then develop an otitis media, for example. Most streptococci can be treated with penicillins, which can also be used during pregnancy.
It is important that many common pain relievers, such as aspirin or ibuprofen, may only be taken in certain phases of pregnancy. Therefore, drug therapy during pregnancy should be discussed with a doctor.
There are still many different infections that can occur during pregnancy. To be better prepared for anything, check out our next topic below: Infections in Pregnancy
What is strep angina?
Streptococcal angina, also known as acute tonsillar angina, is inflammation of the tonsils. The most common causative agent of this disease is the Streptococcus pyogenes.
This angina is particularly common in children between the ages of 3 and 14. The streptococci are transmitted from one person to the next via a droplet infection.
The symptoms of streptococcal angina can vary widely and asymptomatic courses are also possible. Fever, headache, cough and sore throat may occur. The swollen tonsils make speech sound lumpy and swallowing can be painful. Nausea or abdominal pain are also possible.
The diagnosis is mainly made by examining the pharynx. The palatine tonsils appear enlarged and reddened in the event of an infection. White specks can also be deposited on them.
A throat swab or an examination of the bacteria is possible, but not always necessary. A blood test is not carried out routinely as part of the initial diagnosis.
Prognosis: acute tonsillitis comes quickly, but also goes away quickly. If the therapy is carried out properly, the chances of recovery are very high.
For more information on how long strep angina lasts, see: Duration of acute tonsillitis
Therapy mainly consists of adequate fluid intake and the administration of painkillers. If streptococcal infection is detected, the antibiotic penicillin V can be given.
For more information on this topic, see: Acute tonsillar angia
What is strep sepsis
Sepsis is often referred to colloquially as blood poisoning. Formally, this is not entirely correct. A new definition describes sepsis as a disturbed function of the organ systems due to a presumed infection, which can be life-threatening.
Streptococci are the third most common cause of sepsis. Streptococcal sepsis typically begins with an infection, which then leads to pneumonia or soft tissue inflammation, for example. If the immune system does not work properly, an inflammatory reaction occurs throughout the body and sepsis.
If you're more interested in this topic, then check out our next article below: Streptococcal sepsis
Is it possible to vaccinate against streptococci?
You can only vaccinate against pneumococci. Children are vaccinated against pneumococci at 2, 4 and 11-14 months of age. If no complete basic immunization has taken place by the 14th month of life, you can still vaccinate until the 23rd month of life.
Premature babies are also vaccinated at the age of 3 months, for a total of 4 times. In addition, a further vaccination against pneumococci is recommended by the STIKO for people aged 60 and over.
The following topic could also be of interest to you: Why you should vaccinate
These tests for strep throat are there
If streptococcal angina is suspected, a streptococcal A rapid test can be carried out. To do this, take a little throat secretion and mix it with a given liquid. The mixture is then applied to a test kit. The result can be read after just a few minutes.
Read more about this under: Streptococcus rapid test
The test is highly specific but not very sensitive. This means that a negative result cannot rule out an infection. If the result is positive, however, one can be relatively sure of an infection.
A bacterial culture can be set up for any strep infection. For this you need a little secretion from the affected body region. This can be throat or vaginal secretions or wound fluid if a wound infection is suspected.
This secretion is then specially processed and cultivated. When cultivating, after a while you can see which bacteria are in the test substance. One disadvantage is that this method can be relatively tedious.
Recommendations from the editorial team
Further general information could also be of interest to you:
- bacteria
- Illustration of bacteria
- Germs
- Bacteria in the blood - how dangerous is it?
- Bacteria in the intestine - which of them are contagious?










.jpg)





.jpg)